Tóm tắt kiến thức trọng tâm Sinh học 12 ôn thi THPT Quốc gia năm 2021
Nhằm mục đích giúp học sinh có thêm tài liệu ôn tập cho kì thi THPT Quốc gia năm 2021 môn Sinh học, VietJack biên soạn loạt bài Tóm tắt kiến thức trọng tâm Sinh học 12 tổng kết lại những nội dung lý thuyết quan trọng của chương trình Sinh học lớp 12. Hi vọng tài liệu này như là một cuốn sổ tay lý thuyết môn Sinh học giúp học sinh chuẩn bị tốt cho kì thi Đại học sắp tới.
Tóm tắt kiến thức trọng tâm Sinh học 12 ôn thi THPT Quốc gia năm 2021
Nhằm mục đích giúp học sinh có thêm tài liệu ôn tập cho kì thi THPT Quốc gia năm 2021 môn Sinh học, VietJack biên soạn loạt bài Tóm tắt kiến thức trọng tâm Sinh học 12 tổng kết lại những nội dung lý thuyết quan trọng của chương trình Sinh học lớp 12. Hi vọng tài liệu này như là một cuốn sổ tay lý thuyết môn Sinh học giúp học sinh chuẩn bị tốt cho kì thi Đại học sắp tới.
Lý thuyết trọng tâm Cơ chế di truyền và biến dị
- Gen là gì ? Lý thuyết về Gen hay, chi tiết
- Phiên mã, dịch mã là gì
- Điều hòa gen là gì? Cơ chế điều hòa gen ở sinh vật nhân sơ
- Đột biến gen là gì? Nguyên nhân, cơ chế đột biến gen
- Nhiễm sắc thể là gì ? Hình thái, cấu trúc nhiễm sắc thể
- Đột biến nhiễm sắc thể là gì
- Đột biến số lượng nhiễm sắc thể là gì? Cơ chế phát sinh, hậu quả, ý nghĩa
- Công thức về Cơ chế di truyền và biến dị
Lý thuyết trọng tâm Tính quy luật của hiện tượng di truyền
- Quy luật phân li là gì? Nội dung, ý nghĩa ?
- Quy luật phân li độc lập là gì? Nội dung, ý nghĩa ?
- Liên kết gen là gì ? Ý nghĩa
- Hoán vị gen là gì ? Ý nghĩa
- Di truyền liên kết giới tính và di truyền ngoài nhân là gì
- Tương tác gen và tác động đa hiệu của gen là gì
- Ảnh hưởng của môi trường lên sự biểu hiện của gen là gì
- Công thức về tính quy luật của hiện tượng di truyền hay, chi tiết
Lý thuyết trọng tâm Di truyền học quần thể
Lý thuyết trọng tâm Ứng dụng di truyền học
- Chọn giống dựa trên nguồn biến dị tổ hợp
- Tạo giống bằng phương pháp gây đột biến và công nghệ tế bào
- Tạo giống nhờ công nghệ gen
Lý thuyết trọng tâm Di truyền học ở người
Lý thuyết trọng tâm Bằng chứng và cơ chế tiến hóa
- Lý thuyết Bằng chứng tiến hóa hay, chi tiết
- Các học thuyết tiến hóa cổ điển và hiện đại hay, chi tiết
- Loài sinh học là gì ? Lý thyết Loài hay, chi tiết
- Lý thuyết Các nhân tố tiến hóa hay, chi tiết
- Quá trình hình thành đặc điểm thích nghi hay, chi tiết
- Quá trình hình thành loài mới hay, chi tiết
- Tiến hóa lớn là gì
Lý thuyết trọng tâm Sự phát sinh và phát triển của sự sống trên trái đất
- Lý thuyết Nguồn gốc của sự sống
- Sự phát triển của sinh giới qua các các đại địa chất hay, chi tiết
- Lý thuyết Sự phát sinh loài người
Lý thuyết trọng tâm Cá thể và quần thể sinh vật
- Lý thuyết Môi trường sống và các nhân tố sinh thái
- Quần thể sinh vật là gì ? Đặc điểm cơ bản của quan hệ trong quần thể
- Các kiểu, nguyên nhân gây biến động số lượng cá thể của quần thể sinh vật
Lý thuyết trọng tâm Quần xã sinh vật
- Quần xã sinh vật là gì ? Đặc trưng cơ bản của quần xã sinh vật
- Diễn thế sinh thái là gì? Nguyên nhân gây diễn thế sinh thái
Lý thuyết trọng tâm Hệ sinh thái sinh quyển và bảo vệ môi trường
Gen là gì ? Lý thuyết về Gen
1. Định nghĩa
- Gen là một đoạn của phân tử ADN mang thông tin mã hoá một chuỗi pôlipeptit hay một phân tử ARN
- Mỗi gen cấu trúc mã hoá prôtêin bao gồm 3 vùng trình tự
+ Vùng điều hoà: nằm ở đầu 3’ của mạch mã gốc của gen, có trình tự nuclêôtit đặc biệt giúp ARN pôlimeraza có thể nhận biết và liên kết để khởi động quá trình phiên mã đồng thời cũng điều hoà hoạt động phiên mã.
+ Vùng mã hoá: mang thông tin mã hoá các axit amin. Các gen ở sinh vật nhân sơ có vùng mã hoá liên tục trong khi phần lớn các gen ở sinh vật nhân thực có vùng mã hoá không liên tục (gen phân mảnh).
+ Vùng kết thúc: nằm ở đầu 5’ của mạch mang mã gốc, mang tín hiệu kết thúc phiên mã.
2. Mã di truyền
- Mã di truyền là mã bộ ba
- Trong số 64 bộ ba mã hoá có 3 bộ ba không mã hoá cho axit amin nào là: UAA, UAG, và UGA. Đây là các bộ
- Bộ ba AUG là mã mở đầu, đồng thời mã hoá cho axit amin mêtiônin (ở sinh vật nhân sơ là foocmin mêtiônin).
- Các đặc điểm của mã di truyền:
+ Tính phổ biến: tất cả các loài đều có chung một bộ mã di truyền, trừ một vài ngoại lệ.
+ Mã di truyền được đọc từ 1 điểm theo từng bộ 3 mà không gối lên nhau.
+ Tính đặc hiệu: một bộ ba chỉ mã hoá cho 1 axit amin.
+ Tính thoái hoá: nhiều bộ ba khác nhau cùng mã hoá cho 1 axit amin.
3. Quá trình nhân đôi ADN
- Thời gian, địa điểm: Diễn ra trong nhân tế bào vào kì trung gian
- Nguyên tắc: Quá trình tự nhân đôi của ADN dựa hoàn toàn trên NTBS và nguyên tắc bán bảo toàn (giữ lại một nửa)
* Quá trình:
- Bước 1: Tháo xoắn ADN mẹ
- Nhờ các enzim tháo xoắn, 2 mạch đơn của phân tử ADN mẹ tách nhau dần tạo nên chạc chữ Y và để lộ ra 2 mạch khuôn, trong đó một mạch có đâu 3’-OH, còn mạch kia có đầu 5’-P.
- Bước 2: Enzim ADN-pôlimeraza lần lượt liên kết các nuclêôtit tự do từ môi trường nội bào với các nuclêôtit trên mỗi mạch khuôn theo nguyên tắc bổ sung. Vì enzim ADN-pôlimeraza chỉ tổng hợp mạch mới theo chiều 5’ → 3’ nên trên mạch khuôn có đầu 3’ thì mạch bổ sung được tổng hợp liên tục theo chiều 5’→ 3’cùng chiều với chiều tháo xoắn, trên mạch khuôn có đầu 5’ thì mạch bổ sung được tổng hợp ngắt quãng tạo nên các đoạn ngắn gọi là đoạn Okazaki cũng theo chiều 5’→ 3’ ngược chiều với chiều tháo xoắn, sau đó các đoạn này được nối lại với nhau nhờ enzim nối ADN - ligaza
Bước 3 : Hai phân tử mới được tạo thành
Mạch mới tổng hợp đến đâu thì 2 mạch đơn ( một mạch được tổng hợp và một mạch cũ của phân tử ban đầu) đóng xoắn lại với nhau tạo thành hai phân tử ADN con đầu
Kết thúc quá trình nhân đôi: Hai phân tử ADN con được tạo thành có cấu trúc giống hệt nhau và giống ADN mẹ ban
Phiên mã, dịch mã là gì ?
1. Phiên mã
a. Cấu trúc:
- ARN trong tế bào được phân thành 3 loại chủ yếu là: mARN (ARN mang thông tin), tARN (ARN vận chuyển) và rARN (ARN tạo ribôxôm)
- mARN (ARN thông tin) có cấu tạo mạch thẳng được dùng làm khuôn cho quá trình dịch mã. Đầu 5’ có trình tự đặc biệt là nơi bám cho ribôxôm. Sau khi tổng hợp xong prôtêin, mARN thường bị enzim phân huỷ.
- tARN (ARN vận chuyển) có cấu trúc mạch đơn nhưng cuộn lại tao các thuỳ, trong đó có 1 thuỳ chứa bộ ba đối mã (anti-codon). tARN có chức năng vận chuyển axit amin đến ribôxôm và đóng vai trò đọc mã trên mARN thành trình tự axit amin.
- rARN (ARN ribôxôm) là thành phần cấu tạo nên ribôxôm cùng prôtêin.
b. Quá trình tổng hợp ARN (Quá trình phiên mã/sao mã)
- Thời gian, địa điểm: Quá trình tổng hợp ARN diễn ra trong nhân tế bào, tại các NST ở kì trung gian đang ở dạng sợi mảnh
- Nguyên tắc: Quá trình tổng hợp ARN dựa theo NTBS, trong đó A trên mạch gốc liên kết với U, T trên mạch gốc liên kết với A, G liên kết với X và X liên kết với G.
* Quá trình tổng hợp:
Bước 1. Khởi đầu:
Enzym ARN pôlimeraza bám vào vùng điều hoà làm gen tháo xoắn để lộ ra mạch gốc có chiều 3’→ 5’ và bắt đầu tổng hợp mARN tại vị trí đặc hiệu.
- Bước 2. Kéo dài chuỗi ARN:
Enzym ARN pôlimeraza trượt dọc theo mạch gốc trên gen có chiều 3’ → 5’ và các nuclêôtit trong môi trường nội bào liên kết với các nucluotit trên mạch gốc theo nguyên tắc bổ sung:
Agốc - Umôi trường
Tgốc - Amôi trường
Ggốc – Xmôi trường
Xgốc – Gmôi trường
- Bước 3. Kết thúc:
Khi enzym di chuyển đến cuối gen, gặp tín hiệu kết thúc thì quá trình phiên mã dừng lại, phân tử ARN: được giải phóng. Vùng nào trên gen vừa phiên mã xong thì 2 mạch đơn đóng xoắn ngay lại.
Ở tế bào nhân sơ, mARN sau phiên mã được sử dụng trực tiếp cho quá trình dịch mã. Còn ở tế bào nhân thực, sau khi phiên mã, mARN phải được cắt bỏ các đoan intron, nối exon lại với nhau để thành mARN trưởng thành.
* Kết quả, ý nghĩa:
- Từ gen ban đầu tạo ra ARN tham gia quá trình tổng hợp prôtêin ngoài nhân tế bào.
2. Dịch mã
a. Hoạt hoá axit amin
- Dưới tác động của 1 số enzim, các a.a tự do trong môi trường nội bào được hoạt hoá và liên kết với tARN tương ứng → phức hợp a.a – tARN.
b. Tổng hợp chuỗi pôlipeptit
- Bước 1. Mở đầu
+ Tiểu đơn vị bé của ribôxôm gắn với mARN ở vị trí nhận biết đặc hiệu (gần bộ ba mở đầu) và di chuyển đến bộ ba mở đầu (AUG).
+ aa mở đầu - tARN tiến vào bộ ba mở đầu (đối mã của nó – UAX- khớp với mã mở đầu – AUG – trên mARN theo nguyên tắc bổ sung), sau đó tiểu phần lớn gắn vào tạo ribôxôm hoàn chỉnh.
- Bước 2. Kéo dài chuỗi pôlipeptit
+ aa1 - tARN tiến vào ribôxôm (đối mã của nó khớp với mã thứ nhất trên mARN theo nguyên tắc bổ sung), một liên kết peptit được hình thành giữa axit amin mở đầu với axit amin thứ nhất.
+ Ribôxôm chuyển dịch sang bộ ba thứ 2, tARN vận chuyển axit amin mở đầu được giải phóng. Tiếp theo, aa2 - tARN tiến vào ribôxôm (đối mã của nó khớp với bộ ba thứ hai trên mARN theo nguyên tắc bổ sung), hình thành liên kết peptit giữa axit amin thứ hai và axit amin thứ nhất.
+ Ribôxôm chuyển dịch đến bộ ba thứ ba, tARN vận chuyển axit amin mở đầu được giải phóng.
Quá trình cứ tiếp tục như vậy đến bộ ba tiếp giáp với bộ ba kết thúc của phân tử mARN. Như vậy, chuỗi pôlipeptit liên tục được kéo dài.
- Bước 3. Kết thúc
+ Khi ribôxôm chuyển dịch sang bộ ba kết thúc (UAA, UAG, UGA) thì quá trình dịch mã ngừng lại, 2 tiểu phần của ribôxôm tách nhau ra. Một enzim đặc hiệu loại bỏ axit amin mở đầu và giải phóng chuỗi pôlipeptit, quá trình dịch mã hoàn tất.
Trong dịch mã, mARN thường không gắn với từng riboxom riêng rẽ mà đồng thời gắn với một nhóm ribôxôm (pôliribôxôm hay pôlixôm) giúp tăng hiệu suất tổng hợp prôtêin.
Điều hòa gen là gì ? Cơ chế điều hòa gen ở sinh vật nhân sơ
1. Khái niệm
- Điều hoà hoạt động của gen là điều hoà lượng sản phẩm của gen được tạo ra.
- Điều hoà hoạt động của gen nhằm đảm bảo cho hoạt động sống của tế bào phù hợp với điều kiện môi trường cũng như hoạt động sống của cơ thể.
- Quá trình điều hoà hoạt động của gen diễn ra ở nhiều cấp độ khác nhau như điều hoà phiên mã, điều hoà dịch mã, điều hoà sau phiên mã
2. Cơ chế điều hoà gen ở sinh vật nhân sơ
- Ôpêrôn là nhóm các gen có liên quan về chức năng thường phân bố liền nhau thành từng cụm , có chung một cơ chế điều hòa.
a) Mô hình cấu trúc ôpêrôn Lac
- Mô hình cấu trúc ôpêrôn Lac được phát hiện bởi 2 nhà khoa học Pháp là F. Jacôp và J. Mônô ở vi khuẩn E. coli
- Cấu trúc của ôpêrôn Lac
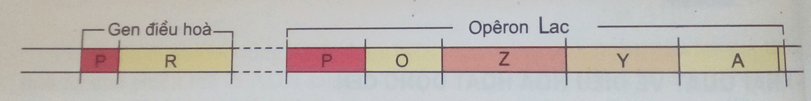
b) Ôpêrôn Lac bao gồm:
+ 3 gen Z, Y, A: là các gen cấu trúc quy định tổng hợp enzim tham gia vào quá trình phân giải đường lactôzơ trong môi trường.
+ O (operator): vùng vận hành, nơi gắn prôtêin ức chế quá trình phiên mã.
+ P (promotor): vùng khởi động, nơi ARN pôlimeraza gắn vào để khởi động phiên mã.
+ Trước ôpêrôn có một gen điều hoà R. Gen R khi hoạt động sẽ tổng hợp prôtêin ức chế. Prôtêin này bám vào vùng vận hành O ngăn cản quá trình phiên mã.
- Hoạt động của ôpêrôn Lac
+ Khi môi trường không có lactôzơ: gen điều hoà tổng hợp prôtêin ức chế. Prôtêin này liên kết với vùng vận hành ngăn cản hoạt động của ARN pôlimeraza làm cho các gen cấu trúc không hoạt động
+ Khi môi trường có lactôzơ: một số phân tử lactôzơ liên kết với prôtêin ức chế làm biến đổi cấu hình không gian của loại pr ô têin thay đổi nên không thể liên kết được với vùng vận hành nữa. Do vậy ARN pôlimeraza có thể thực hiện hoạt động phiên mã các gen Z, Y, A. Sau khi lact ô zơ được phân giải hết, thì prôtêin ức chế được giải phóng và lại liên kết trở lại với vùng vận hành quá trình phiên mã bị dừng lại.
Đột biến gen là gì ? Nguyên nhân, cơ chế đột biến gen
1. Khái niệm
- Đột biến gen là những biến đổi trong cấu trúc của gen. Những biến đổi này thường liên quan đến một (đột biến điểm) hoặc một vài cặp gen.
- Trong tự nhiên, các gen đều có thể bị đột biến nhưng với tần số rất thấp (10-6 – 10-4).
- Tần số đột biến gen có thể thay đổi tuỳ thuộc vào tác nhân đột biến và độ bền của gen.
- Cá thể mang đột biến biểu hiện thành kiểu hình được gọi là thể đột biến.
- Trong điều kiện nhân tạo, có thể chủ động sử dụng các tác nhân gây đột biến để tăng tần số đột biến và định hướng vào 1 gen cụ thể để tạo những sản phẩm tốt phục vụ sản xuất và đời sống
2. Các dạng đột biến
a. Đột biến thay thế một cặp nuclêôtit
Một cặp nuclêôtit này được thay thế bởi 1 cặo nuclêôtit khác có thể làm thay đổi trình tự axit amin trong chuỗi pôlipeptit và thay đổi chức năng của prôtêin.
b. Đột biến thêm hoặc mất một cặp nuclêôtit.
Một cặp nuclêôtit bị mất đi hoặc được thêm vào sẽ làm dịch chuyển khung đọc dẫn đến đọc sai mã di truyền từ vị trí xảy ra đột biến dẫn đến thay đổi trình tự axit amin trong chuỗi pôlipeptit và làm thay đổi chức năng của prôtêin.
3. Nguyên nhân và cơ chế
a. Nguyên nhân
- Do những sai sót ngẫu nhiên phát sinh trong quá trình tự nhân đôi của ADN.
- Tác động của các tác nhân vật lí, hoá học và sinh học trong môi trường.
- Đột biến có thể phát sinh trong điều kiện tự nhiên hoặc do con người tạo ra.
b. Cơ chế phát sinh
- Sự kết cặp không đúng trong tái bản ADN: Các bazơ nitơ thường tồn tại 2 dạng cấu trúc: dạng thường và dạng hiếm. Các dạng hiếm có những vị trí liên kết hiđrô bị thay đổi dẫn đến chúng có thể bắt cặp sai khi tái bản gây đột biến.
- Tác động của các tác nhân gây đột biến:
+ Tác nhân vật lí: tia UV có thể làm cho 2 Timin cạnh nhau trên cùng 1 mạch liên kết với nhau dẫn đến đột biến gen.
+ Tác nhân hoá học: 5BU là chất đồng đẳng của Timin, gây đột biến thay thế cặp A – T bằng cặp G – X.
+ Tác nhân sinh học: dưới tác động của một số loại virut như virut Hecpet, virut viêm gan B, … cũng có thể gây đột biến
4. Hậu quả và ý nghĩa
a. Hậu quả:
- Đa số đột biến gen gây hại, một số có thể có lợi hoặc trung tính đối với thể đột biến.
- Ở mức độ phân tử, đa số đột biến điểm thường là đột biến trung tính.
- Mức độ gây hại của alen đột biến phụ thuộc vào tổ hợp gen cũng như điều kiện môi trường.
b. Vai trò:
- Cung cấp nguyên liệu cho quá trình tiến hóa. Dù tần số đột biến của 1 gen rất thấp nhưng trong tế bào có rất nhiều gen và số lượng cá thể trong quần thể cũng rất lớn, do đó số lượng gen đột biến được tạo ra ở mỗi thế hệ rất lớn - nguồn biến dị di truyền cho quá trình tiến hoá.
- Đột biến gen cũng cung cấp nguyên liệu cho quá trình chọn giống. Người ta chủ động sử dụng các tác nhân đột biến để tạo ra các giống mới